
Ιστορική αναδρομή: Έχει προταθεί ότι τα φλοιϊκά νευρωνικά συστήματα της γλώσσας αποτελούν εξέλιξη των κινητικών συστημάτων του εγκεφαλικού φλοιού και συγκεκριμένα των μετωποβρεγματικών συστημάτων που ενεργοποιούνται κατά τη διάρκεια της παρατήρησης πράξεων. Αν και προηγούμενες έρευνες υποδεικνύουν την ύπαρξη κοινών φλοιϊκών συστημάτων για την πράξη – ή την παρατήρηςή της – και τη γλώσσα, δεν έχει ποτέ τεθεί το ερώτημα κατά πόσον η γλωσσική επεξεργασία των οπτικών ερεθισμάτων προκύπτει αποκλειστικά μέσα στην υποομάδα των μετωποβρεγματικών περιοχών που ενεργοποιούνται κατά τη διάρκεια της παρατήρησης της πράξης. Αν κάτι τέτοιο πράγματι παρατηρηθεί θα ενισχύσει ιδιαίτερα την υπόθεση πως η γλώσσα προέκυψε ως φυσική εξέλιξη των μετωποβρεγματικών συστημάτων τα οποία εμπλέκονται στην αντιστοίχηση της εκτέλεσης μιας πράξης με την παρατήρηση της ίδιας πράξης. Μεθοδολογία/Κύρια Ευρήματα: Χρησιμοποιήσαμε λειτουργική Απεικόνιση Μαγνητικού Συντονισμού (λειτουργική μαγνητική τομογραφία, fMRI) σε άτομα που κοιτούσαν βίντεο αλληλεπιδράσεων χεριού με αντικείμενα, ερεθίσματα ελέγχου με φωτογραφίες των ίδιων αντικειμένων, ενώ παράλληλα εκτελούσαν γλωσσικά (εννοιολογικά και φωνολογικά) και αντιληπτικά έργα. Τα ερεθίσματα ήσαν ίδια για τα γλωσσικά και τα αντιληπτικά έργα, ώστε οι τυχόν διαφορετικές ενεργοποιήσεις να σχετίζονται με τις εκάστοτε απαιτήσεις του ίδιου του έργου. Σύμφωνα με τα αποτελέσματα, τα γλωσσικά έργα ενεργοποίησαν τις κατώτερες μετωπιαίες περιοχές αριστερά, οι οποίες αποτελούν υποσύνολο ενός ευρύτερου αμφίπλευρου μετωποβρεγματικού δικτύου που ενεργοποιήθηκε κατά τη διάρκεια της αντίληψης πράξεων. Καμία μεμονωμένη φλοιϊκή περιοχή δεν έδειξε αποκλειστική – ή έστω εντονότερη – ενεργοποίηση στα γλωσσικά έργα, σε σύγκριση με την αντίστοιχη ενεργοποίησή της στα έργα αντίληψης πράξεων. Συμπεράσματα: Τα αποτελέσματα αυτά δείχνουν ότι τα γλωσσικά έργα όχι μόνο έχουν κοινές νευρωνικές αναπαραστάσεις με την παρατήρηση πράξεων αλλά ουσιαστικά ενεργοποιούν μια υποομάδα του δικτύου παρατήρησης πράξεων, όταν έχουν χρησιμοποιηθεί ίδια ερεθίσματα. Τα ευρήματά μας αποτελούν ισχυρό επιχείρημα υπέρ της εξελικτικής υπόθεσης, σύμφωνα με την οποία συστήματα του μετωποβρεγματικού λοβού που ευθύνονται για την αντιστοίχηση της εκτέλεσης της πράξης με την παρατήρησή της «συμπροσαρμόστηκαν» για τη γλώσσα, μια διεργασία γνωστή ως «εξαρμογή».1
Εισαγωγή
Από τότε που παρατηρήθηκαν για πρώτη φορά στους μακάκους οι κατοπτρικοί νευρώνες –κύτταρα που ενεργοποιούνται τόσο όταν ο πίθηκος εκτελεί στοχοκατευθυνόμενες πράξεις [1], όσο και όταν το ζώο παρατηρεί τις πράξεις κάποιου άλλου– πιθανολογήθηκε ότι τα κύτταρα αυτά έχουν διαδραματίσει ενδεχομένως κάποιο ρόλο στην εξέλιξη της γλώσσας [2-4]. Τα θεωρητικά επιχειρήματα ήταν ουσιαστικά δύο: πρώτον, οι κατοπτρικοί νευρώνες είχαν αρχικά ανακαλυφθεί σε μια περιοχή του εγκεφάλου των μακάκων (περιοχή F5) που φέρεται ως ομόλογη με το μείζον γλωσσικό κέντρο του ανθρώπινου εγκεφάλου, την περιοχή Broca. Δεύτερον, οι κατοπτρικοί νευρώνες φαίνεται να διευκολύνουν την ισοτιμία μεταξύ του πομπού και του δέκτη ενός μηνύματος, μια ισοτιμία που εγκαθιδρύει οτιδήποτε έχει σημασία στην επικοινωνία [5,6]. Πρόσφατα, διάφορα εργαστήρια διερεύνησαν την ύπαρξη κοινών νευρωνικών συστημάτων ανάμεσα στη γλώσσα και την κινητική συμπεριφορά εν γένει, αλλά και ειδικότερα ανάμεσα στη γλώσσα και τις προκινητικές περιοχές που ανταποκρίνονται στην παρατήρηση πράξεων (διαθέτοντας έτσι κατοπτρικές ιδιότητες). Το σύνολο των προηγούμενων ερευνών έχει δείξει κοινούς νευρωνικούς μηχανισμούς – που αφορούν τόσο τους χάρτες ενεργοποίησης [7-9] όσο και την τροποποίηση της νευρωνικής διεγερσιμότητας [10-13] μεταξύ περιοχών της γλώσσας και της κινητικής εν γένει συμπεριφοράς, αλλά και, ειδικότερα, της παρατήρησης πράξεων. Δεν έχει, ωστόσο, διερευνηθεί πειραματικά μέχρι σήμερα ο βαθμός στον οποίο οι νευρωνικοί μηχανισμοί της γλωσσικής επεξεργασίας των οπτικών ερεθισμάτων είναι ανεξάρτητοι από το μετωποβρεγματικό σύστημα των κατοπτρικών νευρώνων (ένα εντελώς διαφορετικό ζήτημα σχετίζεται προφανώς με την αντίληψη του λόγου και τον ανώτερο κροταφικό φλοιό αλλά δεν θα διερευνηθεί εδώ). Οι πειραματικές συνθήκες στις προηγούμενες έρευνες διέφεραν πολύ μεταξύ τους (π.χ. κινητικά έργα ή έργα παρατήρησης πράξεων από τη μια και ανάγνωση λέξεων ή προτάσεων από την άλλη), καθιστώντας με αυτόν τον τρόπο εξαιρετικά δυσχερή την ερμηνεία των διαφορετικών ενεργοποιήσεων μεταξύ γλώσσας και πράξης (ή παρατήρησης της πράξης). Η παρούσα μελέτη υιοθέτησε έναν σχεδιασμό στον οποίο τα πειραματικά ερεθίσματα ήταν ίδια, ενώ διέφεραν οι οδηγίες των έργων, στοχεύοντας είτε σε γλωσσικές λειτουργίες είτε στην αντίληψη της πράξης. Χρησιμοποιώντας αυτόν τον σχεδιασμό πιστεύουμε ότι είμαστε σε θέση να εξετάσουμε το βαθμό στον οποίο η γλωσσική επεξεργασία των οπτικών ερεθισμάτων, που αναπαριστούν πράξεις και αντικείμενα, και οι ανθρώπινες μετωποβρεγματικές περιοχές, που ανταποκρίνονται στην παρατήρηση της πράξης, αλληλεπικαλύπτονται ή διαφέρουν. Το ερώτημα αυτό θεωρούμε ότι είναι σχετικό με την υπόθεση ότι οι κατοπτρικοί νευρώνες έπαιξαν σημαντικό ρόλο στην εξέλιξη της γλώσσας. Αν και οι εξελικτικές υποθέσεις δεν μπορούν να αποδειχθούν πλήρως στο εργαστήριο, προτείνουμε ότι η υπόθεση για τον ρόλο των κατοπτρικών νευρώνων στην εξέλιξη της γλώσσας οδηγεί σε μια σχετικά απλή και κατεξοχήν ελεγχόμενη πρόβλεψη. Εάν οι κατοπτρικοί νευρώνες επιλέχθηκαν αρχικά για την παρατήρηση πράξεων (και πιθανότατα για την κατανόησή τους) και στη συνέχεια συμπροσαρμόστηκαν για τη γλώσσα –μια διεργασία κοινώς γνωστή ως «εξαρμογή» [14]–, θα περίμενε κανείς ότι κατά τη διάρκεια επεξεργασίας ταυτόσημων οπτικών ερεθισμάτων ένα γλωσσικό έργο θα πρέπει να ενεργοποιεί μια υποπεριοχή ή και όλες τις περιοχές που εμπλέκονται στο έργο αντίληψης πράξεων, ενώ δεν θα πρέπει να ενεργοποιείται καμία επιπλέον περιοχή. Εάν τα γλωσσικά έργα ενεργοποιούσαν επιπλέον περιοχές, οι πρόσθετες αυτές περιοχές θα ήταν πιθανότατα αποκλειστικά γλωσσικές στη φύση τους και δεν θα μπορούσαν να αποτελούν προϊόν εξέλιξης των κατοπτρικών νευρώνων.
Μέθοδος
Μελετήθηκαν 14 υγιείς δεξιόχειρες, όλοι τους με μητρική γλώσσα την αγγλική (μέσος όρος ηλικίας 25,1±2,6 έτη, πέντε άνδρες). Η μελέτη εγκρίθηκε από την αρμόδια Επιτροπή του Πανεπιστημίου της Καλιφόρνιας στο Λος Άντζελες και όλοι οι συμμετέχοντες έδωσαν γραπτή συγκατάθεση για τη συμμετοχή τους στη μελέτη.
Διαδικασία και ερεθίσματα
30 χρηστικά αντικείμενα της καθημερινής ζωής προβλήθηκαν είτε ως στατικές εικόνες είτε μέσω βίντεο, όπου ένα χέρι χειριζόταν με συνηθισμένο τρόπο το αντικείμενο (π.χ. φωτογραφία ενός κουδουνιού – βίντεο ενός χεριού που χτυπά το κουδούνι). Οι δύο αυτές ομάδες ερεθισμάτων (φωτογραφίες, ταινίες βίντεο) χρησιμοποιήθηκαν σε τρία έργα (βλ. Σχήμα 1) τα οποία κλήθηκαν να εκτελέσουν οι συμμετέχοντες:
Μια σύνοψη του πειραματικού σχεδιασμού δίνεται στο Σχήμα 1. Οι απαντήσεις των υποκειμένων δίνονταν πατώντας τα πλήκτρα μιας συμβατής με την MRI συσκευής, με το δείκτη ή το μεσαίο δάκτυλο του δεξιού χεριού. Σε όλα τα έργα, σε 30% των ερεθισμάτων, η σωστή απάντηση ήταν «ναι».
Πριν από το κύριο πείραμα απεικόνισης, μια μεγαλύτερη ομάδα ερεθισμάτων δόθηκε σε μια διαφορετική ομάδα 10 υγιών ατόμων με μητρική γλώσσα την αγγλική. Από τους συμμετέχοντες στην προκαταρκτική αυτή συμπεριφορική μελέτη ζητήθηκε να αξιολογήσουν αν τα αντικείμενα που τους προβάλλονταν ήταν εργαλεία που χρησιμοποιούνται για κατασκευές και ποια λέξη θα περιέγραφε καλύτερα το κάθε αντικείμενο. Έτσι, για τα 30 ερεθίσματα που χρησιμοποιήθηκαν στο πείραμα της απεικόνισης εγκεφάλου, το σύνολο των 10 αυτών μαρτύρων είχε συμφωνήσει τόσο στο όνομα του αντικειμένου όσο και στο ότι το αντικείμενο είναι εργαλείο.

Μελέτη fMRI
Κάθε πειραματικό έργο με λειτουργική μαγνητική τομογραφία περιλάμβανε έξι διακριτές πειραματικές ομάδες (blocks). Η σειρά της προβολής των έξι αυτών ομάδων εξισορροπήθηκε μεταξύ των συμμετεχόντων βάσει ενός ψευδοτυχαίου σχεδιασμού. Η διάρκεια προβολής κάθε ομάδας ήταν 24 δευτερόλεπτα. Σε ενδιάμεσες ομάδες αναφοράς, που διαρκούσαν 20 δευτερόλεπτα, οι συμμετέχοντες έπρεπε να κοιτούν μία οριζόντια γραμμή στο κέντρο της οθόνης (συνθήκη ανάπαυσης).
Επί 1 δευτερόλεπτο πριν από τα διακριτά έργα κάθε πειραματικής ομάδας, προβαλλόταν μια σύντομη πρόταση η οποία δήλωνε αποκλειστικά το είδος του έργου που ακολουθούσε, δεδομένου ότι τα ερεθίσματα εικόνων και βιντεοταινιών ήταν ίδια μεταξύ των έργων.
Πραγματοποιήθηκε απεικόνιση με λειτουργική μαγνητική τομογραφία (fMRI) βασισμένη στην αντίθεση επιπέδου οξυγόνωσης του αίματος (BOLD) (3T Siemens Allegra scanner, Ahmanson-Lovelace Brain Mapping Center, και ένα πηνίο κεφαλής). Η διαδικασία της fMRI περιλάμβανε 2 εικονικές σαρώσεις, τις οποίες ακολούθησαν 600 σαρώσεις ολόκληρου του εγκεφάλου (υπερταχεία ακολουθία, single-shot gradient-refocused echo-planar imaging – EPI) (TR =2.0 s, TE = 25 ms, γωνία επανόρθωσης = 90°, 36 τομές).
Ανάλυση δεδομένων
Τα δεδομένα της fMRI αναλύθηκαν με χρήση του λογισμικού Statistical Parametric Mapping software (SPM2, http://www.fil.ion.ucl.ac.uk /spm/, London, UK). Οι εικονικές σαρώσεις δεν χρησιμοποιήθηκαν. Οι υπόλοιπες σαρώσεις αναδιατάχθηκαν και κανονικοποιήθηκαν σε έναν τυπικό στερεοταξικό χώρο χρησιμοποιώντας τον οδηγό EPI του Montreal Neurological Institute (MNI). Το μέγεθος του ογκοστοιχείου (voxel) ήταν 3x3x3mm. Στη συνέχεια, τα κανονικοποιημένα δεδομένα ομαλοποιήθηκαν με τη χρήση ενός Γκαουσιανού πυρήνα (Gaussian kernel) 8x8x8 mm, προκειμένου να βελτιωθεί η αναλογία σήματος-θορύβου. Η εκτίμηση των επόμενων παραμέτρων έγινε με χρήση κατάλληλου σχεδιασμού που χρησιμοποίησε τη λειτουργία box-car2 σε συνδυασμό με τη λειτουργία της αιμοδυναμικής απόκρισης. Προκειμένου να αποκλειστούν τα σήματα χαμηλής συχνότητας, τα δεδομένα φιλτραρίστηκαν με υψιπερατό (high-pass) φίλτρο (περίοδος διακοπής 128 s). Ο υπολογισμός της παραμέτρου ογκοστοιχείο προς ογκοστοιχείο για τα εξομαλυμένα δεδομένα πραγματοποιήθηκε σύμφωνα με το γενικό γραμμικό μοντέλο. Αρχικά υπολογίστηκε η απλή αντίθεση για κάθε έργο και η σύγκριση μεταξύ των έργων για τον κάθε συμμετέχοντα, όπως λεπτομερώς περιγράφεται παρακάτω. Οι εικόνες αντίθεσης αποτέλεσαν τη βάση για τις συνολικές ενεργοποιήσεις fMRI με χρήση ενός μοντέλου τυχαίων επιδράσεων (σύγκριση μέσων όρων με τη χρήση t-test για ένα δείγμα). Το επίπεδο στατιστικής σημαντικότητας καθορίστηκε για όλες τις συγκρίσεις στο 0,05 με διόρθωση λάθους (τιμή ψευδούς διαπίστωσης, FDR), ενώ το επίπεδο στατιστικής σημαντικότητας για τη δημιουργία εκμαγείων στο 0,05, χωρίς διόρθωση λάθους. Αναφέρονται αποκλειστικά οι συστοιχίες με τουλάχιστον 20 γειτνιάζοντα ογκοστοιχεία.
Αν και ο πειραματικός σχεδιασμός της μελέτης ακολούθησε ένα πλήρως παραγοντικό μοντέλο αλληλεπίδρασης του έργου (αντιληπτικό, εννοιολογικό, φωνολογικό) με το ερέθισμα (βίντεο, εικόνα), η υπό διερεύνηση υπόθεση δεν μπορεί να ελεγχθεί μέσω κύριων επιδράσεων και αλληλεπιδράσεων, αλλά με συγκεκριμένες αντιθέσεις. Πράγματι, το έργο παρατήρησης πράξεων (Β-Αντ) ήταν η μόνη συνθήκη όπου η προσοχή κατευθυνόταν προς το χέρι που χειριζόταν το αντικείμενο, ενώ στα υπόλοιπα πέντε έργα η προσοχή κατευθυνόταν στα αντικείμενα. Τα δεδομένα της fMRI αναλύθηκαν με τον ακόλουθο τρόπο, προκειμένου να διερευνηθεί η λειτουργική σχέση μεταξύ του συστήματος παρατήρησης ενεργειών και περιοχών της γλώσσας:
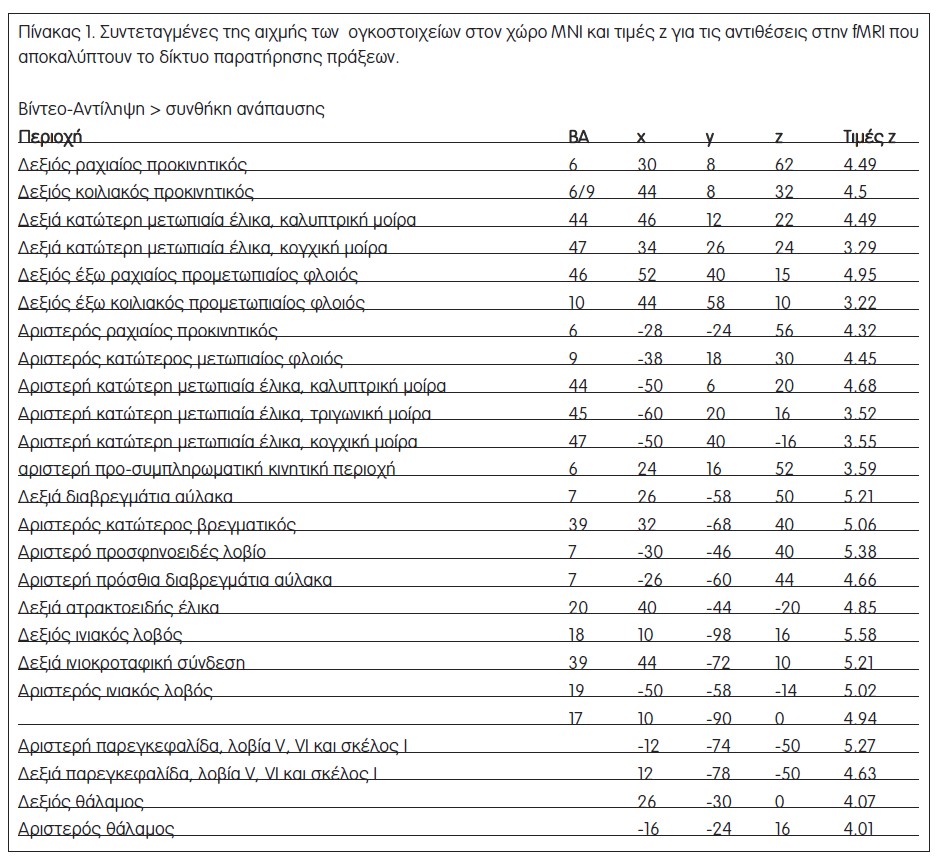
Οι περιοχές που ενεργοποιούνται στην παρατήρηση πράξεων αποκαλύπτονται από την αντίθεση Β-Αντ – συνθήκη ανάπαυσης. Προκειμένου να εκτιμηθεί η διαδικασία λήψης αποφάσεων που εμπλέκεται στη συνθήκη Β-Αντ, συγκρίναμε επίσης μεταξύ τους τις ενεργοποιήσεις κατά τη διάρκεια των συνθηκών Β-Αντ και Ε-Αντ (παρατήρηση αντιληπτικού αντικειμένου). Τα εννοιολογικά και φωνολογικά δίκτυα που εμπλέκονται στα γλωσσικά έργα καταδεικνύονται από τη σύγκριση των συνθηκών Ε-Ενν – ανάπαυση και Ε-Φων – ανάπαυση. Η σύγκριση των γλωσσικών δικτύων με τα δίκτυα παρατήρησης πράξεων αποκαλύφθηκαν από τις συγκρίσεις Ε-Ενν – ανάπαυση και Ε-Φων – ανάπαυση. Η σύγκριση των γλωσσικών δικτύων με τα δίκτυα παρατήρησης πράξεων έγινε σε δύο στάδια: η έκταση των φωνολογικών και εννοιολογικών δικτύων, που σχετίζονται με το δίκτυο παρατήρησης πράξεων, ελέγχθηκε με διαδικασία απόκρυψης, όπου οι ενεργοποιήσεις που παράχθηκαν από τις συγκρίσεις Ε-Φων – ανάπαυση και Ε-Ενν – ανάπαυση καλύφθηκαν αποκλειστικά από τις ενεργοποιήσεις που αποκαλύφθηκαν από την αντίθεση Β-Αντ. Η υπόθεση ότι η γλωσσική επεξεργασία των παρατηρούμενων πράξεων αποτελεί εξέλιξη του μετωποβρεγματικού συστήματος παρατήρησης πράξεων προβλέπει ότι δεν θα υπάρξει ενεργοποίηση συστοιχιών για αυτή την αντίθεση, δηλαδή κανένα μέρος του εννοιολογικού ή του φωνολογικού δικτύου που εξετάζεται εδώ δεν εκτείνεται πέρα από τη δράση του συστήματος παρατήρησης πράξεων. Περαιτέρω συγκρίσεις που εμπλέκονται στις συνθήκες με ίδια ερεθίσματα αποτίμησαν το βαθμό ενεργοποίησης μεταξύ του συστήματος παρατήρησης πράξεων και των γλωσσικών δικτύων (Β-Αντ – (Β-Ενν+Β-Φων) και (Β-Ενν+Β-Φων) – Β-Αντ). Αυτές οι συγκρίσεις σχεδιάστηκαν για μια ποσοτική αντιπαραβολή της δραστηριότητας των περιοχών που ενεργοποιούνται τόσο κατά τη διάρκεια της γλωσσικής επεξεργασίας όσο και κατά τη διάρκεια των έργων αντίληψης πράξεων.
Αποτελέσματα
Επίδοση στο έργο
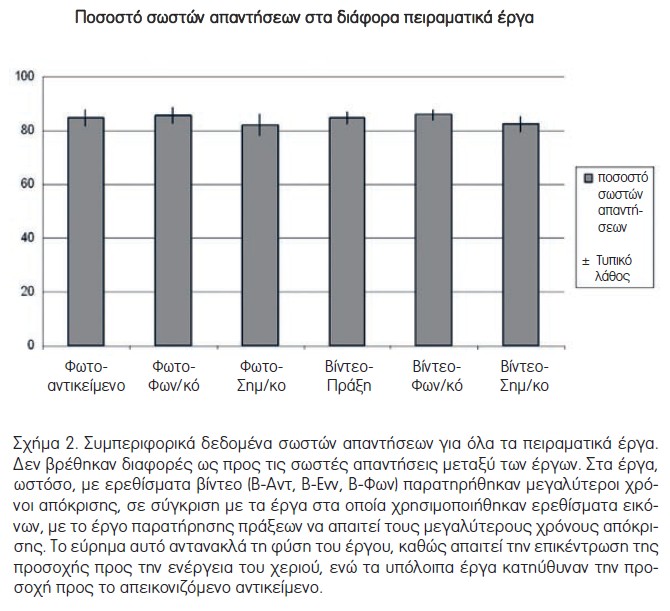
Το μέσο ποσοστό των σωστών απαντήσεων για το σύνολο των έργων ήταν 84.26±1.22%. Τα διορθωμένα ποσοστά για τα έξι διαφορετικά έργα ήταν ως εξής: Ε-Αντ, 84.7±2.9% (S.E.), Ε-Φων, 85.6±3.1%, Ε-Ενν, 82.1±3.7%, Β-Αντ 84.7±2.2%, Β-Φων 85.9±1.9%, Β-Ενν 82.4±2.7% (Σχήμα 2). Αυτά τα ποσοστά σωστών απαντήσεων ενδέχεται να οφείλονται σε δύο παράγοντες. Καταρχάς, στο πείραμα απεικόνισης του εγκεφάλου χρησιμοποιήσαμε έργο κατευθυνόμενης απάντησης και έτσι κάποια από τα λάθη ενδεχομένως οφείλονται σε πολύ γρήγορες αποκρίσεις. Δεύτερον, κάποια από τα ερεθίσματα ενείχαν αντιληπτικές αμφιβολίες, για παράδειγμα μόνο ένα μικρό μέρος του αντικειμένου ήταν μαύρο ή μόνο τέσσερα δάχτυλα κρατούσαν το αντικείμενο. Σύγκριση μέσων όρων των σωστών απαντήσεων για εξομοιωμένα ζεύγη και με χρήση του t-test (με διόρθωση για πολλαπλές συγκρίσεις) δεν έδειξε στατιστικά σημαντικές διαφορές μεταξύ των συνθηκών.
Ο μέσος χρόνος απόκρισης για το σύνολο των συνθηκών ήταν 1284.9±31.4 ms. Ο χρόνος απόκρισης για τις μεμονωμένες συνθήκες και από την έναρξη του ερεθίσματος κυμάνθηκε μεταξύ 1096.2±42.4 ms και 1546.2±74.2 ms (Ε-Ενν: 1103.0±8.6 ms, Ε-Φων: 1193.6±44.3 ms, Ε-Ενν: 1096.2±42.4 ms, Β-Αντ: 1546.2±74.2 ms, Β-Φων 1395.5±52.7 ms, Β-Ενν 1375.0±55.4 ms). Σύγκριση των μέσων όρων των χρόνων απόκρισης για εξομοιωμένα ζεύγη έδειξε στατιστικά σημαντικά μεγαλύτερους χρόνους απόκρισης για το έργο παρατήρησης πράξεων, σε σύγκριση με όλα τα υπόλοιπα έργα, για τα δύο έργα με ερεθίσματα βίντεο (Β-Φων και Β-Ενν), σε σύγκριση με τα τρία έργα που είχαν ως ερεθίσματα εικόνες και για το Ε-Φων σε σύγκριση με το Ε-Ενν. Αυτές οι διαφορές φαίνονται λογικές. Πράγματι το έργο αντίληψης με χρήση βίντεο μπορούσε να λυθεί μόνο τη χρονική στιγμή που το χέρι ακουμπούσε το αντικείμενο. Αντίθετα, η απαραίτητη πληροφορία για την εκτέλεση των υπολοίπων έργων ήταν παρούσα από το πρώτο καρέ του βίντεο.
Αποτελέσματα της λειτουργικής απεικόνισης
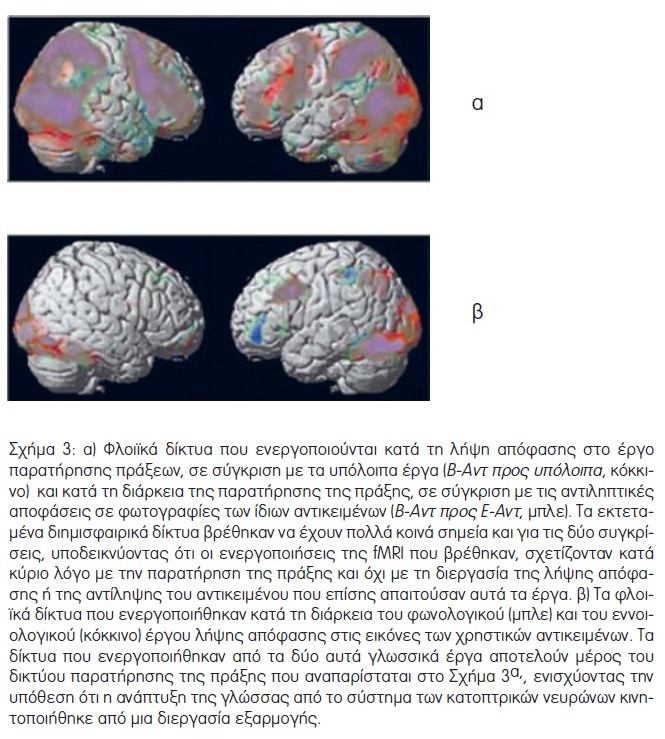
Όπως αναμενόταν, η αντίθεση Β-Αντ προς ανάπαυση αποκάλυψε αμφίπλευρη αύξηση των σημάτων στις εξωταινιωτές οπτικές περιοχές, τις κατώτερες και ανώτερες βρεγματικές περιοχές και εκτεταμένες αμφίπλευρες ημισφαιριακές μετωπιαίες ενεργοποιήσεις, που περιλάμβαναν προκινητικές, κατώτερες μετωπιαίες και προμετωπιαίες (έξω ραχιαίος και έξω κοιλιακός προμετωπιαίος φλοιός) περιοχές (Σχήμα 3α, Πίνακας 1). Αυτό το δίκτυο βρίσκεται σε συμφωνία με το αμφίπλευρο δίκτυο που περιγράφεται σε προηγούμενες μελέτες fMRI που διερεύνησαν την παρατήρηση πράξεων [7,15–18]. Η αντίθεση Β-Αντ – Ε-Αντ, η οποία αφαίρεσε την ενεργοποίηση που οφειλόταν στην αντίληψη του αντικειμένου και τη λήψη αποφάσεων, έδειξε ένα εξαιρετικά παρόμοιο δίκτυο αμφίπλευρων βρεγματικών, προκινητικών και προμετωπιαίων περιοχών (Σχήμα 3α).
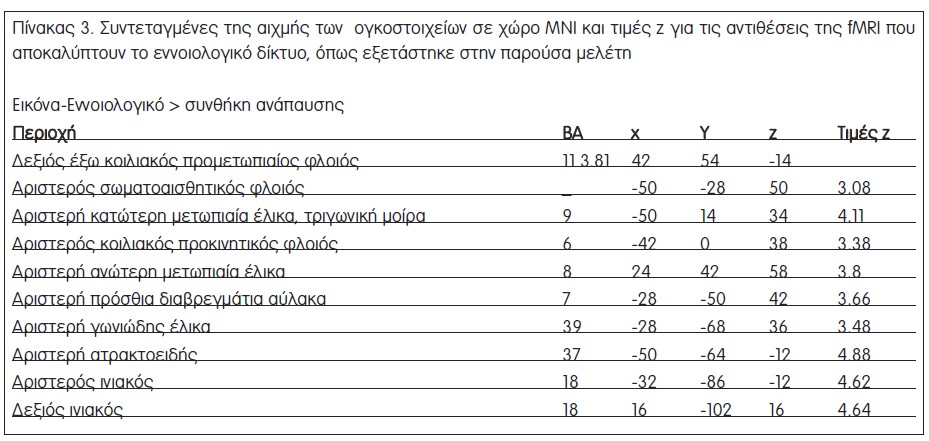
Η αντίθεση Ε-Φων προς ανάπαυση έδειξε κυρίως αριστερή μετωπιαία ενεργοποίηση, συμπεριλαμβανομένων πρόσθιων κατώτερων μετωπιαίων περιοχών και της παρακείμενης μέσης μετωπιαίας έλικας, του κοιλιακού και του έξω ραχιαίου προμετωπιαίου φλοιού, και μια αριστερή ενεργοποίηση της υπερχείλιας έλικας (Σχήμα 3β, Πίνακας 2). Παρόμοια, το εννοιολογικό έργο (Ε-Φων προς ανάπαυση) ενεργοποίησε ένα, κατά κύριο λόγο, αριστερά πλαγιωμένο δίκτυο που περιλαμβάνει μία συστοιχία στην οπίσθια αριστερή μετωπιαία έλικα, η οποία εκτείνεται στην κάτω ραχιαία μετωπιαία έλικα και τον κοιλιακό προκινητικό φλοιό και μια βρεγματική συστοιχία που καλύπτει τη γωνιώδη έλικα (Σχήμα 3β, Πίνακας 3). Αυτές οι περιοχές συμφωνούν με τις βασικές περιοχές που περιγράφονται σε μια πρόσφατη μεταανάλυση μελετών fMRI που ασχολήθηκαν με φωνολογικές και εννοιολογικές επεξεργασίες [19]. Δεν βρέθηκαν σημαντικές κροταφικές ενεργοποιήσεις, βάσει του δεδομένου ουδού που τέθηκε, για κανένα από τα δύο γλωσσικά έργα, ένα αποτέλεσμα που πιθανότατα σχετίζεται με το γεγονός ότι ο πειραματικός σχεδιασμός χρησιμοποίησε ως ερεθίσματα των γλωσσικών έργων φωτογραφίες αντικειμένων αντί για γραπτό κείμενο ή λόγο.
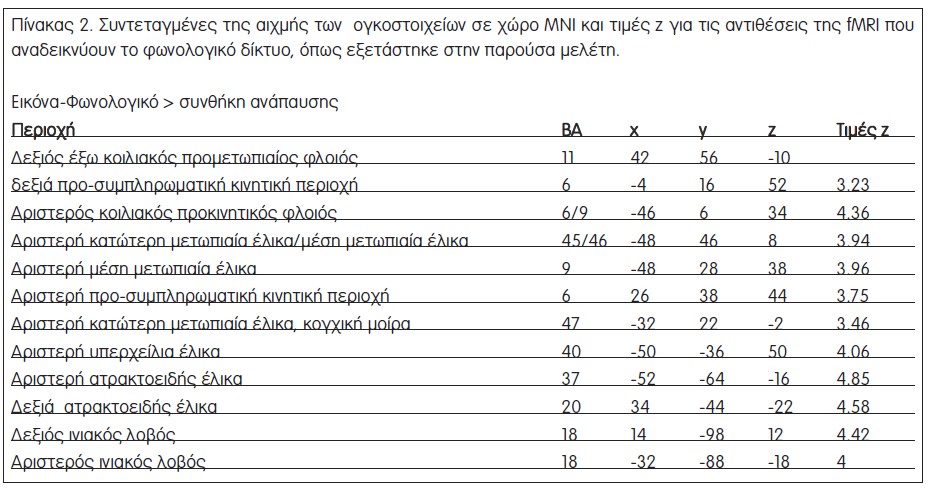
Το δεύτερο βήμα της ανάλυσης των δεδομένων είχε στόχο τη συστηματική σύγκριση των φλοιϊκών δικτύων, που εμπλέκονται στην παρατήρηση της πράξης και στην εννοιολογική/φωνολογική επεξεργασία, προκειμένου να φωτίσει την υπόθεση κοινών εξελικτικά επεξεργασιών από τα δίκτυα της γλώσσας και το σύστημα των κατοπτρικών νευρώνων. Επικάλυψη των ενεργοποιήσεων του φωνολογικού δικτύου (Ε-Φων – ανάπαυση) από τον χάρτη ενεργοποίησης στο έργο αντιληπτικής πράξης (Β-Αντ – ανάπαυση) δεν έδειξε κάποια επιπλέον ενεργοποίηση συστοιχιών. Η αντίστοιχη ανάλυση απόκρυψης για το εννοιολογικό δίκτυο (Ε-Ενν – ανάπαυση) είχε τα ίδια αποτελέσματα. Τα αποτελέσματα αυτά δείχνουν ότι δεν υπήρξαν φλοιϊκές περιοχές που να ενεργοποιήθηκαν αποκλειστικά κατά τη διάρκεια είτε του εννοιολογικού έργου είτε του φωνολογικού έργου, και οι οποίες δεν αποτελούν μέρος του δικτύου που ενεργοποιήθηκε από την αντίληψη πράξεων.
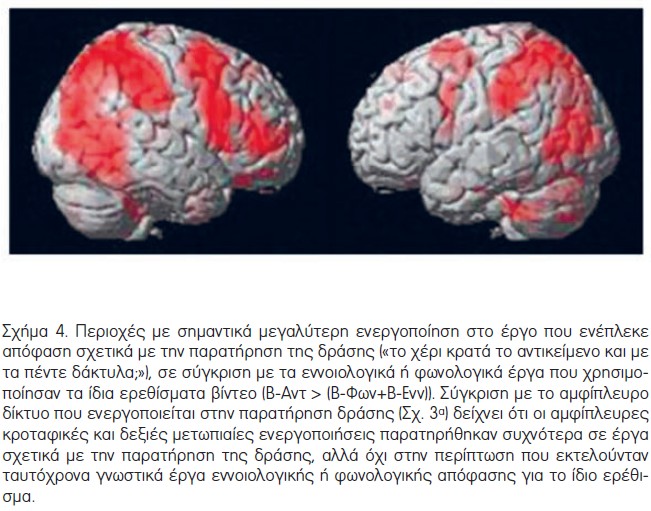
Περαιτέρω, οι στατιστικές συγκρίσεις πραγματοποιήθηκαν με αναφορά στο βαθμό ενεργοποίησης που παρατηρήθηκε κατά τη διάρκεια της φωνολογικής ή της εννοιολογικής επεξεργασίας, σε σύγκριση με την αντίληψη της πράξης. Για αυτή τη σύγκριση, χρησιμοποιήθηκαν οι τρεις συνθήκες με ίδια ερεθίσματα βίντεο (Β-Αντ, Β-Φων και Β-Ενν). Η συγκριτική ανάλυση των ενεργοποιήσεων fMRI, κατά τη διάρκεια αυτών των έργων, [(Β-Ενν+Β-Φων) προς Β-Αντ, Β-Ενν προς Β-Αντ, Β-Φων προς Β-Αντ] δεν έδειξε στατιστικά σημαντική μεγαλύτερη ενεργοποίηση για το εννοιολογικό ή το φωνολογικό έργο, σε σύγκριση με την ενεργοποίηση κατά το έργο αντίληψης της πράξης. Η αντίστροφη σύγκριση (Β-Αντ προς (Β-Ενν+Β-Φων)) κατέδειξε ένα εκτεταμένο αμφίπλευρο βρεγματικό και κυρίως δεξιό μετωπιαίο δίκτυο περιοχών που παρουσίασαν μεγαλύτερη ενεργοποίηση κατά τη διάρκεια αντίληψης της πράξης, σε σύγκριση με τη γλωσσική ανάλυση, δεδομένων των ταυτόσημων ερεθισμάτων βίντεο (Σχήμα 4, Πίνακας 4). Μικρές μόνο συστοιχίες στον αριστερό μετωπιαίο λοβό παρουσίασαν μεγαλύτερη ενεργοποίηση για την αντίληψη της πράξης, σε σύγκριση με την εννοιολογική και φωνολογική επεξεργασία. Αυτό το εύρημα, σε συνδυασμό με τις προηγούμενες αναλύσεις, υποδεικνύει ένα ανάλογο επίπεδο ενεργοποίησης του κατώτερου αριστερού μετωπιαίου φλοιού κατά τη διάρκεια τόσο των έργων αντίληψης της πράξης, όσο και των γλωσσικών έργων.
Συζήτηση
Το πείραμα που παρουσιάστηκε εδώ υιοθέτησε έναν σχεδιασμό που απαιτούσε από τα υποκείμενα να επεξεργαστούν ίδια οπτικά ερεθίσματα, ενώ ταυτόχρονα εκτελούσαν διαφορετικά έργα: ένα έργο αντίληψης πράξεων και δύο «γλωσσικά» έργα, ένα φωνολογικό και ένα εννοιολογικό. Αυτός ο πειραματικός σχεδιασμός μας επιτρέπει να ελέγξουμε κατά πόσον η παρατηρούμενη ρητή γλωσσική επεξεργασία της κατευθυνόμενης προς το αντικείμενο πράξης επιστρατεύει φλοιϊκές περιοχές που δεν εμπλέκονται στην αντίληψη της πράξης και/ή κατά πόσον ενεργοποιεί σε μεγαλύτερο βαθμό μετωποβρεγματικές περιοχές αντίληψης της πράξης. Αυτά τα αποτελέσματα θα υποστήριζαν την υπόθεση κάποιας ανεξαρτησίας της γλωσσικής επεξεργασίας των οπτικών ερεθισμάτων από τις μετωποβρεγματικές περιοχές που ευθύνονται για την αντίληψη της πράξης. Ωστόσο, δεν βρέθηκε κάποια εξειδικευμένη ενεργοποίηση ή αυξημένη δραστηριότητα κάποιας περιοχής κατά τη διάρκεια των δύο γλωσσικών έργων. Υποστηρίζουμε ότι αυτά τα αποτελέσματα είναι συμβατά με την υπόθεση ότι η γλώσσα –τουλάχιστον, όσον αφορά τη γλωσσική επεξεργασία των οπτικών ερεθισμάτων– προέκυψε από τη συμπροσαρμοστική εξέλιξη μετωποβρεγματικών περιοχών που σχετίζονται με την αντίληψη της πράξης, μια διεργασία που είναι γνωστή ως «εξαρμογή».
Αν και προηγούμενες μελέτες αναφέρουν κοινές περιοχές ενεργοποίησης για την πράξη (ή για την παρατήρηση της πράξης) και τη γλώσσα [8,17] –ενεργοποιήσεις που τυπικά ερμηνεύονται ως υποστηρικτικές του πλαισίου της ενσωματωμένης σημασιολογίας (embodied semantics) [20,21]–, αυτές οι προηγούμενες μελέτες δεν θα μπορούσαν να επιχειρηματολογήσουν επάνω στη διακριτή ενεργοποίηση πράξης και γλώσσας, καθώς τα ερεθίσματα που χρησιμοποίησαν διέφεραν ευρέως μεταξύ των έργων πράξης και γλώσσας. Έτσι, το βασικό καινοτόμο εύρημα της μελέτης μας είναι ότι η γλωσσική επεξεργασία οπτικών ερεθισμάτων, η οποία σχετίζεται με την πράξη, προκύπτει σε μια υποομάδα των μετωποβρεγματικών περιοχών που σχετίζονται με την αντίληψη της πράξης.
Μπορεί κανείς να υποστηρίξει ότι η αντίληψη της πράξης συνεπάγεται αυτόματη γλωσσική επεξεργασία και ότι οι κατώτερες μετωπιαίες περιοχές αριστερά, που ενεργοποιούνται με τον ίδιο τρόπο κατά τη διάρκεια της αντίληψης της πράξης και των γλωσσικών έργων, είναι πράγματι αποκλειστικά γλωσσικές στη φύση τους. Αν και αυτό το επιχείρημα είναι λογικά σωστό, είναι απίθανο να αληθεύει. Πράγματι, τα δεδομένα μας δείχνουν ότι ο κατώτερος μετωπιαίος φλοιός έχει επίσης μεγαλύτερη ενεργοποίηση κατά την αντίληψη της πράξης (Β-Αντ), σε σύγκριση με την αντίληψη του αντικειμένου (Ε-Αντ), δυο έργα φαινομενικά παρόμοια όσον αφορά την πιθανότητα αυτόματης γλωσσικής επεξεργασίας, ανόμοια ωστόσο όσον αφορά την ίδια την αντίληψη της πράξης. Έτσι, η «αποπληθωριστική ερμηνεία» (deflationary expla-nation), που επικαλείται αυτόματη γλωσσική επεξεργασία στον αριστερό κατώτερο μετωπιαίο φλοιό για όλα τα έργα, δεν εξηγεί επαρκώς όλα τα ερευνητικά αποτελέσματα που παρουσιάστηκαν εδώ. Περαιτέρω, μια μελέτη εικονικής βλάβης, που χρησιμοποίησε επαναλαμβανόμενο Διακρανιακό Μαγνητικό Ερεθισμό (TMS), έδειξε ότι η παροδική διακοπή της νευρωνικής δραστηριότητας στον κατώτερο αριστερό (και δεξιό) μετωπιαίο φλοιό έχει ως αποτέλεσμα ελλείμματα στη μίμηση, όχι όμως γενικευμένα οπτικοκινητικά ελλείμματα [22]. Τα αποτελέσματα αυτά δύσκολα μπορούν να εναρμονιστούν με μια αμιγώς γλωσσική ιδιότητα του κατώτερου αριστερού μετωπιαίου φλοιού.
Ένας άλλος ισχυρισμός θα μπορούσε να είναι ότι τα αυξανόμενα σήματα στις μετωποβρεγματικές περιοχές οφείλονται αποκλειστικά στις αυξανόμενες απαιτήσεις για προσοχή στο έργο αντίληψης πράξεων, δεδομένων των αυξημένων χρόνων απόκρισης σε αυτό. Θα πρέπει να σημειωθεί, ωστόσο, ότι η αύξηση του χρόνου απόκρισης σε αυτό το έργο ήταν πολύ μικρή, σε σύγκριση με τη συνολική διάρκεια κάθε πειραματικού μέρους του έργου κατά τη διάρκεια της σάρωσης. Ως εκ τούτου, θεωρούμε απίθανο η διαφορά στο χρόνο απόκρισης μεταξύ των έργων να είχε ουσιώδη επίδραση στις ενεργοποιήσεις της λειτουργικής μαγνητικής τομογραφίας. Επιπλέον, το επιχείρημα της «προσοχής» δεν μπορεί να ερμηνεύσει την έλλειψη αυξανόμενων σημάτων στον κατώτερο αριστερό μετωπιαίο φλοιό κατά τη διάρκεια της αντίληψης της πράξης, σε σύγκριση με τα γλωσσικά έργα. Η επιλεκτικότητα της επίδρασης αμφισβητεί έντονα μια εξειδικευμένη επίδραση προσοχής.
Ο πειραματικός σχεδιασμός μας επέτρεψε, επίσης, να συγκρίνουμε τη δραστηριότητα κατά τη διάρκεια τόσο της αντίληψης της πράξης, όσο και της αντίληψης στατικών εικόνων, οι οποίες περιλάμβαναν όλα τα οπτικά στοιχεία των δυναμικών ερεθισμάτων πράξης. Αυτή η σύγκριση έδειξε εγκεφαλική δραστηριότητα, ιδιαίτερα εξειδικευμένη για την παρατήρηση της πράξης και όχι για τα σύνθετα οπτικά στοιχεία που σταθερά συνοδεύουν τις παρατηρούμενες πράξεις. Αυτή η σύγκριση στο πείραμά μας έδειξε ισχυρή αμφίπλευρη ενεργοποίηση των μετωποβρεγματικών περιοχών, καταδεικνύοντας ότι αυτό το εκτεταμένο δίκτυο αφορά ειδικά την παρατήρηση της πράξης. Έτσι, το αποτέλεσμα της συγκεκριμένης σύγκρισης υποστηρίζει την ερμηνεία των «κατοπτρικών νευρώνων», την οποία υποστήριξε μεγάλος αριθμός και προηγούμενων μελετών, οι οποίες έδειξαν παρόμοιες μετωποβρεγματικές ενεργοποιήσεις σε μια ποικιλία πειραματικών συνθηκών [7,18,23–26].
Οι μετωποβρεγματικές περιοχές που συσχετίζονται με την αντίληψη της πράξης κείνται αμφίπλευρα, ενώ τα γλωσσικά έργα επιστράτευσαν αποκλειστικά περιοχές του αριστερού ημισφαιρίου. Αυτή η άρση της αμφίπλευρης ενεργοποίησης για την αντίληψη της πράξης προς ένα κατά κύριο λόγο αριστερά πλαγιωμένο γλωσσικό σύστημα, ενδεχομένως να διευκολύνθηκε, στους ανθρώπους, από την πλαγίωση των «κατοπτρικών» αποκρίσεων σε ήχους πράξεων, όπως έχει δειχθεί με μονοπαλμικό διακρανιακό μαγνητικό ερεθισμό (TMS) [27] και λειτουργική μαγνητική τομογραφία (fMRI) [28].
Καταλήγοντας, όταν τα οπτικά ερεθίσματα που αφορούν πράξεις προς την κατεύθυνση του αντικειμένου γίνονται αντικείμενα αντιληπτικής επεξεργασίας, ενεργοποιούν ένα εκτεταμένο αμφίπλευρο μετωποβρεγματικό δίκτυο. Όταν τα ίδια ερεθίσματα τυγχάνουν γλωσσικής επεξεργασίας, ενεργοποιούν ένα μόνο τμήμα αυτού του δικτύου και καμία επιπλέον περιοχή. Αυτός ο τύπος δραστηριότητας υποστηρίζει την εξελικτική υπόθεση ότι οι νευρωνικοί μηχανισμοί της γλώσσας στον άνθρωπο προέκυψαν φυλογενετικά από παλαιότερους μετωποβρεγματικούς νευρώνες που σχετίζονται με την αντίληψη της πράξης, όπως οι κατοπτρικοί νευρώνες στους μακάκους.
Το περιεχόμενο αυτής της ιστοσελίδας δεν ειναι διαθέσιμο για αντιγραφή.