
Η σύγχρονη νευροεπιστήμη βασίζεται σε έννοιες που προήλθαν από τη νευροανατομία και ανάγονται στη νευρωνική θεωρία του 1888, τότε που ο Ramon y Cajal, χρησιμοποιώντας τη χρώση του Golgi, απέδειξε ότι το νευρικό σύστημα δεν είναι συγκύτιο, αλλά αποτελείται από μεμονωμένους νευρώνες (DeFelipe και Jones, 1988). Η εισαγωγή του ηλεκτρονικού μικροσκοπίου, περίπου 50 χρόνια αργότερα, επέτρεψε να δούμε με υψηλή ευκρίνεια τα χάσματα μεταξύ των νευρώνων και χάρισε στη νευροεπιστήμη ένα ισχυρό εργαλείο διεξοδικού ελέγχου της εσωτερικής δομής της σύναψης, και πιο πρόσφατα, των μεταβολών που υφίσταται στη μάθηση και την αρρώστια.
Η οργάνωση του φλοιού σε στιβάδες και στήλες είναι θεμελιώδης για τη νευρωνική λειτουργία. Ο Brodmann στηρίχτηκε σε διαφορές του σχήματος και της διάταξης των νευρώνων σε φλοιώδεις στιβάδες ώστε να διαιρέσει το φλοιό σε αρχιτεκτονικές περιοχές (Brodmann, 1909). Στην οριζόντια διαστρωμάτωση επικάθεται μια κάθετη οργάνωση, σε στήλες, νευρώνων με κοινά χαρακτηριστικά (Mountcastle et al. 1955). Η διττή αυτή οργάνωση του φλοιού αυξάνει σημαντικά την υπολογιστική του ικανότητα (Grossberg, 1999) και παρέχει τη δομική βάση του διαχωρισμού και της αλληλεπίδρασης μεταξύ εισόδων (inputs) και εξόδων (outputs), του επιτόπιου ανασταλτικού ελέγχου και της παράλληλης και κατανεμημένης επεξεργασίας.
Οι στήλες που είχαν χαρτογραφηθεί σε διάφορες φυσιολογικές μελέτες θεωρήθηκε αρχικά ότι διατρέχουν το φλοιό σε όλο του το βάθος. Ενδείξεις πως αυτό δεν ισχύει σε όλες τις περιπτώσεις εμφανίστηκαν όταν μια απλή ιστοχημική μέθοδος σήμανσης της ενεργότητας της οξειδάσης του κυτοχρώματος κατάτμησε τις στήλες της πρωτοταγούς οπτικής περιοχής των πρωτευόντων, χρωματίζοντας έντονα κάποιες «κηλίδες» στις στιβάδες 2-3, διαχωρίζοντάς τες από περιοχές με χαμηλή σήμανση. Η επαναξιολόγηση των ιδιοτήτων αντίδρασης των νευρώνων στις «κηλίδες» έδειξε πως οι νευρώνες αυτοί ξεχώριζαν επιπλέον και στη φυσιολογία τους (Livingstone και Hubel, 1984).
Μόνο η εφαρμογή ειδικών ιχνηθετών επέτρεψε να γίνουν σαφώς ορατές οι στήλες και έδωσε τη δυνατότητα να κατανοήσουμε την οργάνωσή τους, όπως στην περίπτωση των στηλών της οπτικής επικράτησης στον πρωτοταγή οπτικό φλοιό των πρωτευόντων ή των πεδίων με μορφή βαρελιών στον σωμαισθητικό φλοιό των τρωκτικών. Οι ανωτέρω ιχνηθέτες αντικατέστησαν τις πρωτοποριακές, αλλά δύσκολες και περιορισμένες χαρτογραφικές μεθόδους που χρησιμοποιούσαν την εκτομή-εκφύλιση, και προσέφεραν καινούργιες προοπτικές με εξαιρετικό ενδιαφέρον. Καμιά άλλη τεχνική δεν διαθέτει ανάλογη ισχύ και ευελιξία ώστε να δείχνει ταυτόχρονα το φάσμα εισόδων και εξόδων μικρών ή μεγάλων εγκεφαλικών περιοχών, μια στήλη, μια στιβάδα ή μεμονωμένους νευρώνες.
Χρησιμοποιώντας τέτοιους ιχνηθέτες μάθαμε, για παράδειγμα, πως οι συνδέσεις μεταξύ οιωνδήποτε δύο δομών είναι γενικά αμοιβαίες. Αν και αρχικά δεν ήταν γνωστό από τις λειτουργικές μελέτες, είναι τώρα πλέον σαφές πως οι αμοιβαίες συνδέσεις παίζουν έναν θεμελιώδη ρόλο σε όλα τα νευρωνικά συστήματα, από την απλή αισθητηριακή αντίληψη ως τις σύνθετες νοητικές διεργασίες (για ανασκόπηση βλ. Barbas et al., 2002).
Η «σήμανση» των νευρώνων κατέστησε δυνατή και τη μελέτη των αλληλεπιδράσεων των προμετωπιαίων συνειρμικών περιοχών, οι οποίες δεν ήταν εύκολο να υπαχθούν σε φυσιολογικές μελέτες και παρέμεναν «σιωπηρές» μέχρι πρόσφατα (για ανασκόπηση βλ. Goldman-Rakic, 1996). Οι ιδέες προχώρησαν όταν τα ανατομικά δεδομένα έδειξαν ότι ο προμετωπιαίος λοβός, που για πολύ καιρό εθεωρείτο ως έδρα της νόησης, έχει επίσης και μια μεταιχμιακή συνιστώσα, ένα σύστημα που κλασικά συνδέεται με τα συναισθήματα (Nauta, 1979, Yakovlev, 1948). Ο μεταιχμιακός φλοιός, που βρίσκεται στις έσω και βασικές επιφάνειες, επεκτάθηκε κατά την εξέλιξη των πρωτευόντων παράλληλα με τους συνειρμικούς φλοιούς και διατηρεί ισχυρούς δεσμούς με εκείνους, συνδέοντας αδιάσπαστα περιοχές που σχετίζονται με τη νόηση και με τα συναισθήματα (για ανασκόπηση βλ. Barbas et al., 2002).
Το στοιχείο αυτό θέτει σε αμφισβήτηση την κλασική ιδέα του Πλάτωνα σύμφωνα με την οποία οι σκέψεις και οι συγκινήσεις είναι ολωσδιόλου ξεχωριστά. Στην πραγματικότητα, η αποσύνδεση οδών που συνδέονται με τη νόηση και με τα συναισθήματα βρίσκεται ίσως στην καρδιά ψυχιατρικών νοσημάτων που χαρακτηρίζονται από την ανικανότητα να συνδεθεί το κατάλληλο συναίσθημα με μια δεδομένη κατάσταση. Μια τέτοια παθολογία προσβάλλει δυσανάλογα τις μεταιχμιακές περιοχές και, κατ’ επέκταση, την ανάδρομη επικοινωνία, την επικρατούσα διάταξη των προβολών των μεταιχμιακών περιοχών (Barbas et al., 2002).
Παρά την εμπειρική και εννοιολογική της συμβολή, η νευροανατομία συχνά υποτιμάται όσον αφορά στη δυνατότητά της να κατανοήσει τη νευρωνική λειτουργία. Η αντίληψη αυτή πηγάζει από μια θεμελιώδη παρεξήγηση σύμφωνα με την οποία οι πληροφορίες που αντλούνται από «νεκρούς ιστούς» είναι περιορισμένες, όπως ένας συνάδελφος φυσιολόγος υπαινίχθηκε κάποτε. Άλλαξε κάπως τη γνώμη του τυχαία όταν, εξετάζοντας μαζί ιστολογικές τομές του εγκεφάλου ενός εκ των πιθήκων του, είδαμε ότι μεγάλο μέρος του πλάγιου γονατώδους πυρήνα από τη μια πλευρά είχε εκφυλιστεί. Παραδόξως, η μερική τύφλωση του πιθήκου είχε διαφύγει της προσοχής κατά τη φάση της εκπαίδευσης της συμπεριφοράς και στις φυσιολογικές καταγραφές, συναγόταν όμως τώρα από ένα πακέτο ιστολογικών τομών.
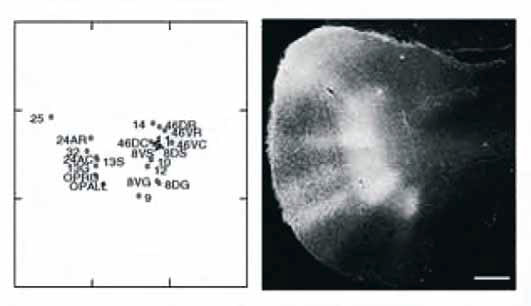
Ωστόσο, πέραν αυτού του είδους των αντιλήψεων, προβλήματα υπάρχουν στη νευροανατομία, τα κυριότερα από τα οποία είναι επίπονοι κατάλογοι όρων, διαφορές ανάμεσα στους χάρτες που δημιουργούν σύγχυση και αρχιτεκτονικά σύνορα που δεν γίνονται εύκολα αντιληπτά από τους περισσότερους. Μπορούν αυτά τα προβλήματα να λυθούν; Πιστεύω πως ναι. Ας πάρουμε για παράδειγμα τις παραλλαγές των χαρτών. Διαθέτουμε τώρα ένα πλήθος μοριακών δεικτών οι οποίοι κατανέμονται διαφορετικά στον φλοιό, δίνοντας τη δυνατότητα κατασκευής ποσοτικών διαγραμμάτων και αντικειμενικού χαρακτηρισμού των αρχιτεκτονικών περιοχών.
Στην εικόνα 1 (αριστερά) παρουσιάζεται ένα τέτοιο παράδειγμα, όπου μια δεκάδα αρχιτεκτονικών χαρακτηριστικών που αξιολογήθηκαν ποσοτικά αντιμετωπίζονται ταυτόχρονα με τη χρήση πολυδιάστατης ανάλυσης. Οι προμετωπιαίες μεταιχμιακές περιοχές συγκεντρώθηκαν αριστερά και οι ευστιβαδικές (eulaminate) δεξιά, διότι διαφέρουν δομικά. Οι αντικειμενικές προσεγγίσεις καταργούν την ανάγκη στήριξης σε υποκειμενικές αναλύσεις και μπορούν να λύσουν το πρόβλημα των παραλλαγών των χαρτών στη βιβλιογραφία.
Υπάρχει ανάγκη αξιόπιστων χαρτών, σήμερα περισσότερο από ποτέ, ώστε να εντοπιστεί η δραστηριότητα και να ερμηνευθούν οι λειτουργικές απεικονιστικές μελέτες στους ανθρώπους. Εντούτοις, η αξία των χαρτών ξεπερνά την ανάγκη τοπογραφικού εντοπισμού διότι η δομή επηρεάζει τη διάταξη των συνδέσεων. Έτσι, συστηματικές παραλλαγές αρχιτεκτονικής αποτελούν το υπόστρωμα της βαθμιδωτής στιβαδωτής διάταξης των φλοιο-φλοιϊκών συνδέσεων (βλ. Barbas et al., 2002). Οι διατάξεις της διαμόρφωσης των συνδέσεων έχουν ερμηνευθεί κατά ποικίλους τρόπους, ότι αντανακλούν δηλαδή, είτε την κατεύθυνση της επεξεργασίας στις αισθητηριακές περιοχές, είτε την απόσταση μεταξύ των συνδεδεμένων περιοχών. Ωστόσο, ακριβώς όπως οι στήλες δεν αποτελούν ιδιαίτερο χαρακτηριστικό των αισθητηριακών περιοχών αλλά αντιπροσωπεύουν θεμελιώδεις οργανωτικές μονάδες του φλοιού (Εικόνα 1, δεξιά), έτσι, και οι συστηματικές παραλλαγές της φλοιώδους δομής μπορεί να χρησιμεύσουν για να εξηγήσουν και να προείπουν, εξίσου καλά, τη μορφή των συνδέσεων στις αισθητηριακές, τις συνειρμικές και τις μεταιχμιακές περιοχές του φλοιού.
Μια συχνά διατυπωμένη κριτική σε βάρος της νευροανατομίας βρίσκεται στο γεγονός ότι είναι περιγραφική. Αλλά η ακριβής, ποσοτική περιγραφή είναι το sine qua non για τις επιστήμες της ζωής, είτε πρόκειται για σεσημασμένα συναπτικά κομβία, είτε για τις ιδιότητες εκφόρτισης των νευρώνων ή την έκφραση των γονιδίων. Ο Δαρβίνος τη χρησιμοποίησε και το ίδιο έκανε ο Ramon y Cajal και οι χαρτογράφοι των ανθρώπινων γονιδίων. Οι ενδελεχείς περιγραφές του Δαρβίνου για τους σπίνους, τα οστρακόδερμα και τους σχηματισμούς των βράχων ήσαν απαραίτητες για την επαγωγική διαδικασία που γέννησε τη θεωρία της εξέλιξης. Τα ποσοτικά, εμπειρικά δεδομένα παρέχουν την ευκαιρία αποκάλυψης σχέσεων, εξαγωγής αρχών, εκτέλεσης υπολογισμών, προτυποποίησης και πρόβλεψης της φυσιολογικής λειτουργίας και της δυσλειτουργίας στη νόσο. Και εκεί ακριβώς βρίσκεται το πάθος και η πρόκληση της νευροανατομίας που, ενισχυμένη με ποσοτικά εργαλεία και αναλύσεις, βρίσκεται στην καρδιά της νευροεπιστήμης.
Το περιεχόμενο αυτής της ιστοσελίδας δεν ειναι διαθέσιμο για αντιγραφή.